THE EVOLUTION OF ANIMAL SOUND-MAKING

WHERE DID the voice come from? How does it work? What is it for? And how does a creature develop one? I am fascinated by these questions, and by animal sound-making in general. In my academic life, I’ve been lucky to work with some amazing scientists to try and answer them.
During my undergraduate years at Yale University, I worked with Richard Prum to study the musicality of birdsong. Bird songs often display musical features such as pitch and rhythm. We wondered whether the ordering of their “notes” also has a musical quality. In an online survey, we presented with human listeners with two versions of songs from twenty different species: one with the notes in original order, and one with them scrambled in time. On average, our listeners rated the original bird songs more musical than the scrambled ones. We then thought through different hypotheses for the origins of avian musicality, including honest signaling, perceptual bias, and arbitrary aesthetic coevolution. You can read the paper in Frontiers in Biology.
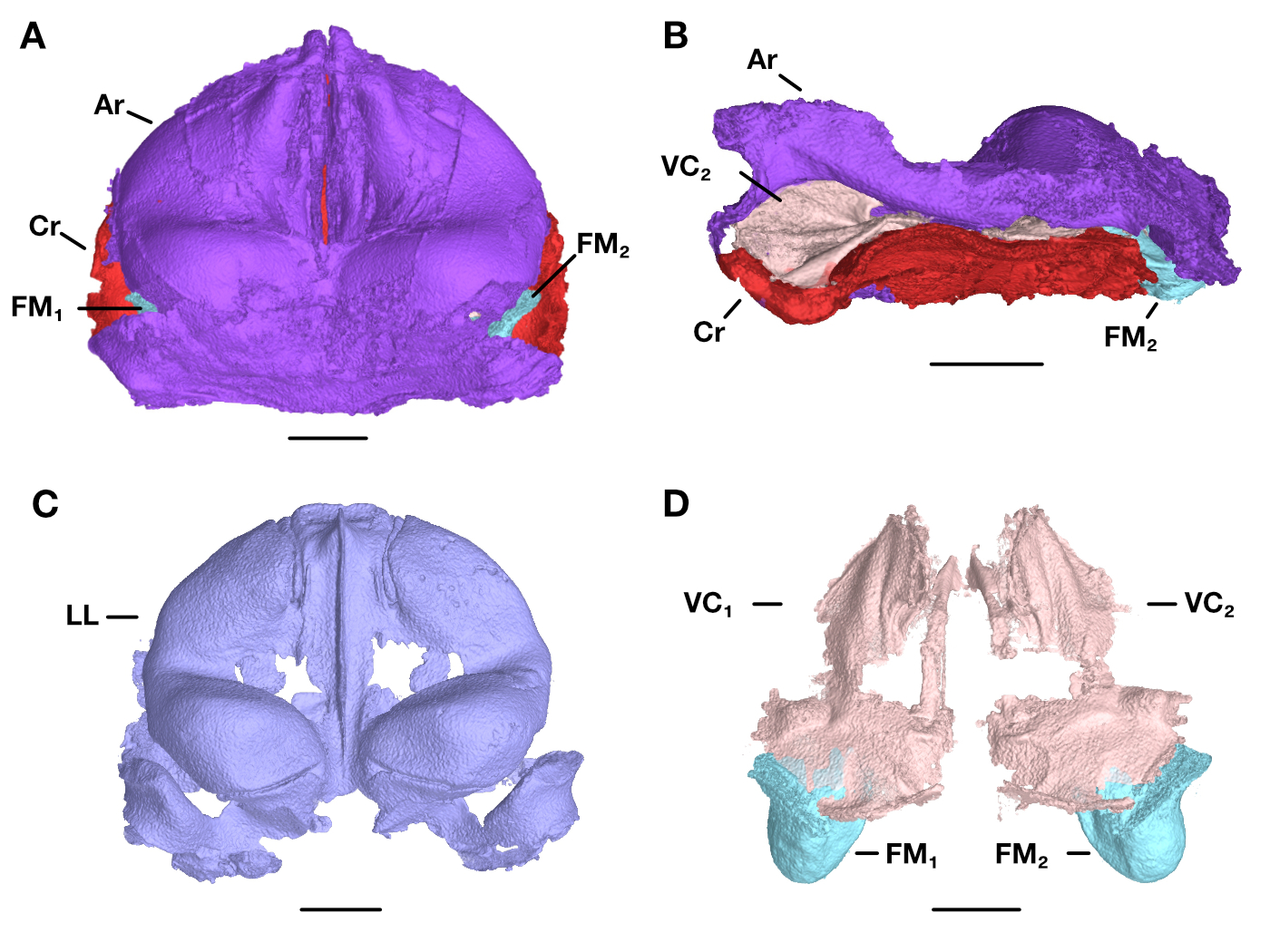
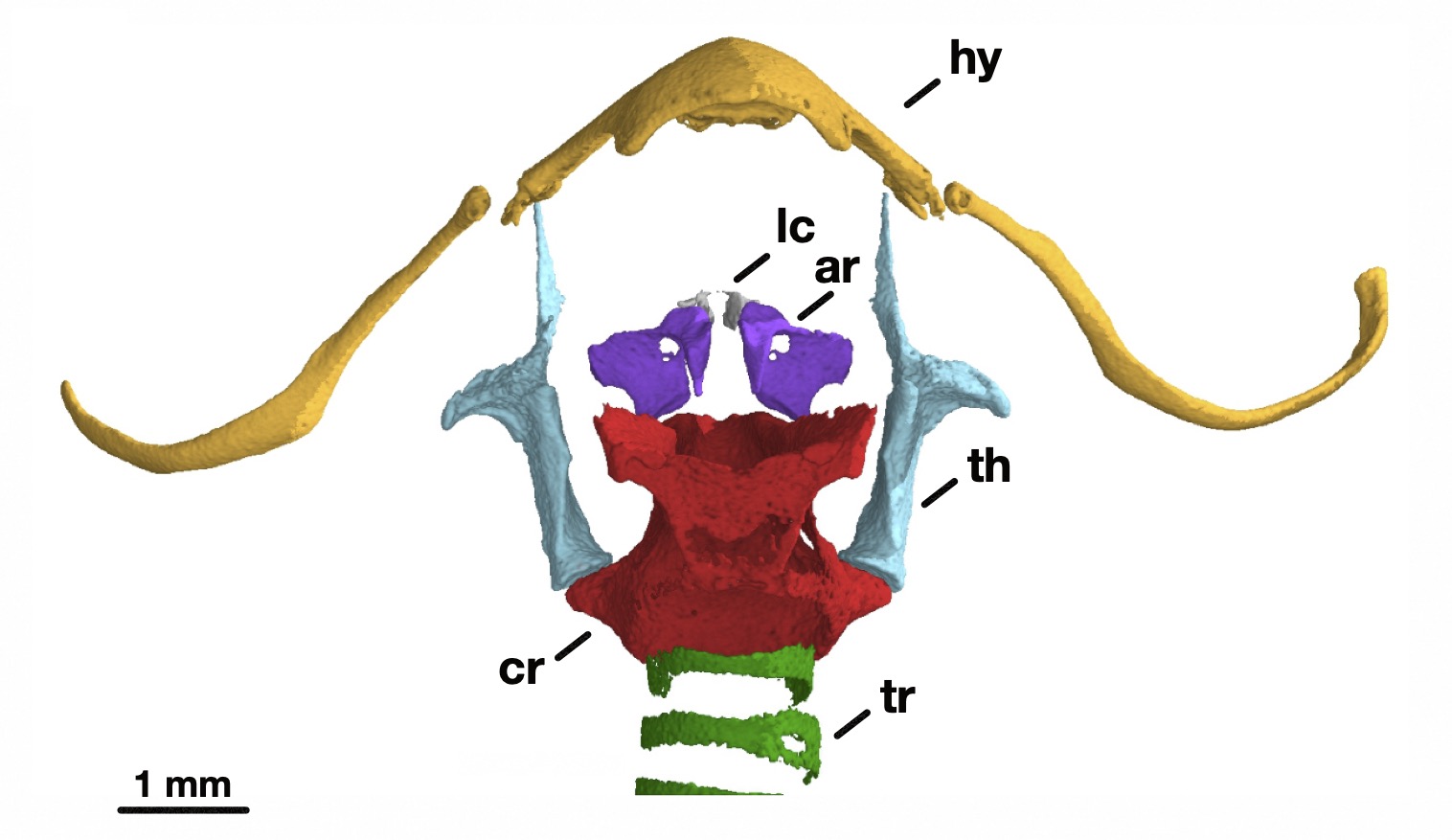
In my PhD work at the University of Texas at Austin, I collaborated with Julia Clarke, Mike Ryan, and Mirjam Knörnschild to study the evolution of the voice in frogs, bats, and humans. In the first chapter of my dissertation, we looked at development of the larynx (voice box) in the túngara frog, which lives in the forests and grasslands of the neotropics. Male túngaras attract females by calling, either alone or in choruses. Calls don’t just attract females, however—they’re also a beacon for frog-eating bats and blood-sucking flies. Males start appearing in choruses and have well-defined vocal cords by the time they’re about 19 mm long, but they don’t begin calling until about 24 mm. Is this because the larynx isn’t fully mature until the latter point, or is it because the calls of small-bodied adults wouldn’t be attractive enough to outweigh the risks of producing them? We approached this question by “tuning” mathematical models of the túngara vocal system with anatomical measurements from CT scans of frogs of different ages. Our results showed that male túngara frogs could likely vocalize during the “silent phase” in their development, suggesting that their muteness is voluntary and not physically enforced. The study, which, like the rest of my dissertation chapters, is currently being prepared for publication, helps to show what happens to anatomy, development, and behavior when natural and sexual selection interact.
My second chapter looked at another group of remarkable animal vocalists: neotropical bats. Some bat species are capable of vocal learning, and their echolocation calls can achieve extremely high pitches (fundamental frequencies up to 250 kHz), short durations (down to 0.2 ms), fast repetitions (up to 220 calls/s), and great intensities (up to 140 dB at 10 cm). Studies of the anatomical underpinnings of the bat voice—the vocal folds, membranes, and tract—have thus far focused on single species. We used advanced imaging techniques such as micro- and diceCT to compare vocal morphology and allometry (how traits scale with body size) across eleven bat species spanning six families. To put these findings in context, we assembled the largest-yet dataset of mammalian fundamental frequency, vocal fold length, and anatomical specializations. Our findings suggest that echolocation fundamental frequency and vocal fold and membrane length are largely determined by body size. We also observed that patterns of laryngeal cartilage mineralization are more shaped by phylogeny (evolutionary history) than the acoustic demands of specific call types. Finally, we failed to find a systematic relationship between larynx morphology, call structure, and vocal learning ability. Overall, our results suggested that strong physical and phylogenetic constraints act on the form and acoustic output of the bat larynx, despite its specializations.
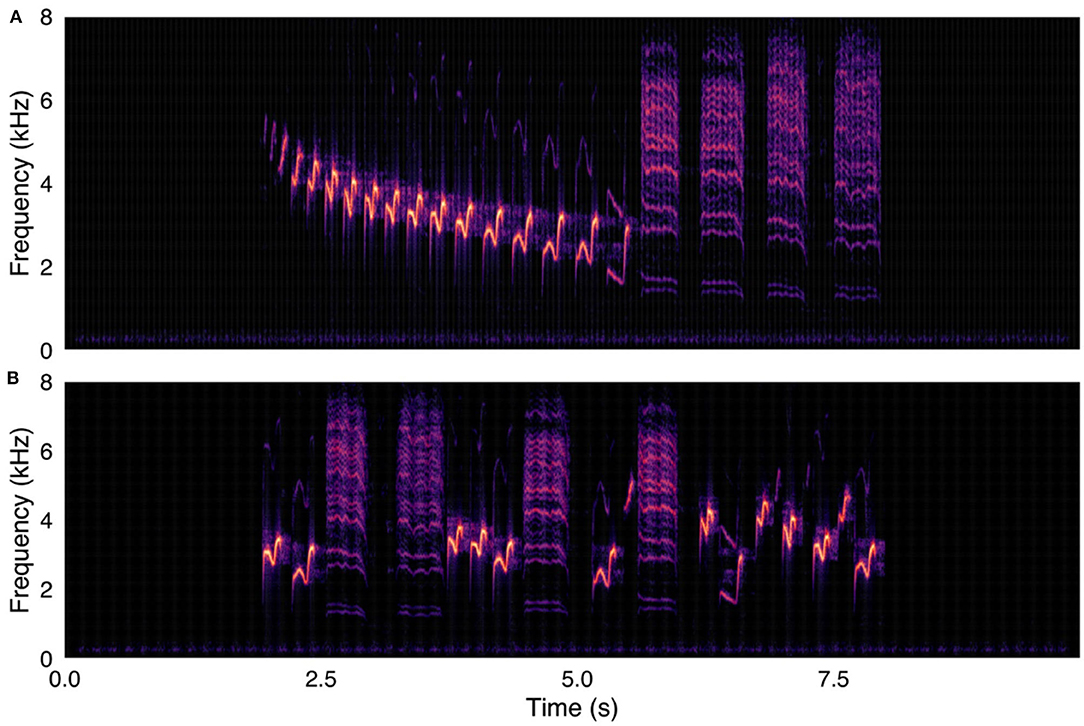
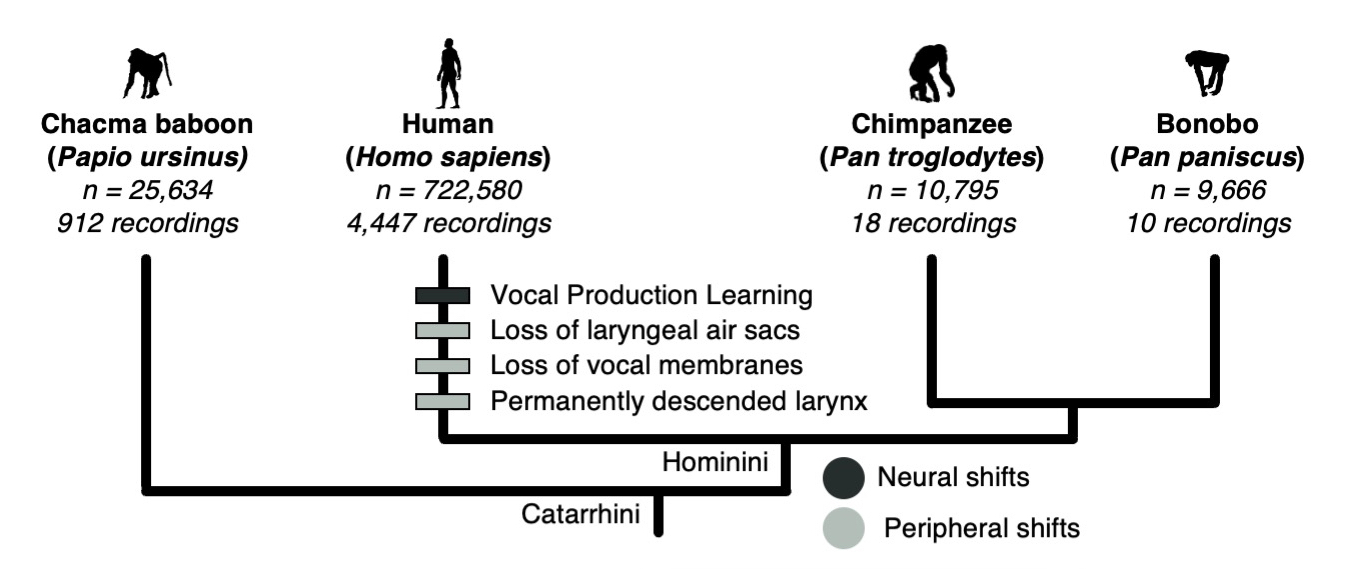
My last chapter was concerned with the evolution of human speech. The human larynx, compared to those of closely related primates, lies deeper in the throat and lacks vocal membranes and air sacs. These shifts are usually analyzed in terms of their acoustic effects on vowel-like vocalizations, since the evolution of spoken language was long thought to require an expansion of vocal range driven by modifications to the vocal tract. However, vowels are just one type of phoneme, and speech is just one class of human utterance. To understand the reasons for these shifts in human vocal anatomy, a broader bioacoustic comparison was needed. Specifically, we needed to compare the range of sound qualities used in human speech to that of other forms of human vocalization and in the repertoires of our close primate relatives. In this study, we measured the amount of acoustic space occupied by a large, cross-cultural sample of human speech, non-linguistic, and musical vocalizations along with the calls of chimpanzees, bonobos, and chacma baboons. We used acoustic markers called Mel-frequency cepstral coefficients to create an acoustic space depicting the sonic features of over 750,000 brief vocal segments. Speech and song turned out to occupy significantly less acoustic-feature volume than human non-linguistic vocalizations. In addition, the volumes of speech and song were not statistically different from those of non-human primates. Our results suggested that while the anatomical shifts we see in humans may have led to an elaboration of non-linguistic utterances, learned vocalizations use a surprisingly small fraction of human acoustic space. Therefore, speech was probably not enabled by an anatomy-driven expansion of human vocal repertoire—more likely, it arose after we acquired vocal learning.
You can download my academic CV here.
PUBLICATIONS


